细胞骨架是细胞主要的机械结构,包含不同的蛋白生物聚合物。它是一个复杂且动态的网络结构。在真核生物中,它由三个主要成分组成,肌动蛋白微丝,中间纤维及微管。这三种纤维都可以根据细胞的要求,通过聚合和分解来快速生长和收缩(Fletcher and Mullins, 2010)。
微管由α及β微管蛋白以二聚体形式组成。它们聚合形成微管蛋白原纤维,最终结合,构成中空微管。微管是细胞骨架的重要组成部分,它支撑了真核生物细胞的形态和结构。
此外,它们也参与了细胞分裂,胞内转运和细胞运动的过程(Goodson and Jonasson, 2018)。
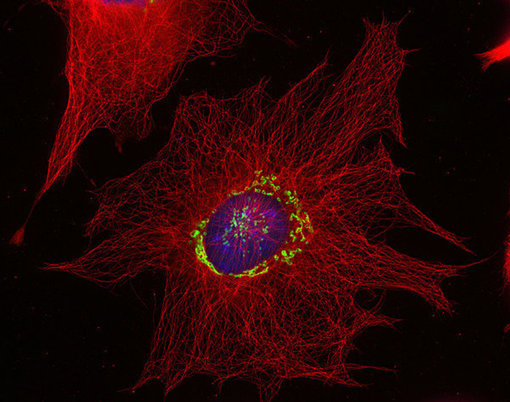
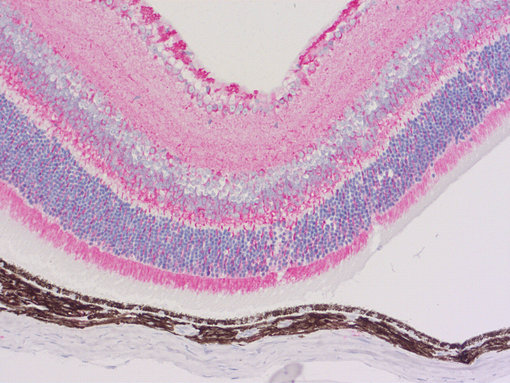
图1:使用抗α-微管蛋白 (cat. no. 302 008, 稀释比例 1 : 500; 红色) 和豚鼠抗巨蛋白(cat. no. 263 005, 稀释比例 1 : 500; 绿色)抗体,对由甲醇固定的小鼠纤维母细胞进行间接免疫染色。细胞核由DAPI染色可见为蓝色。
图2:使用鼠抗α-微管蛋白 (cat. no. 302 211, 稀释比例 1 : 1000; AP-红色)抗体,对由PFA固定,石蜡包埋的小鼠眼切片进行间接免疫染色。细胞核由苏木精染色可见为蓝色。
作为管家基因,微管蛋白在所有细胞类型中都有大量表达。因此在蛋白免疫印迹实验中,它们经常被用作内参对照。它们也通常用标记蛋白来可视化免疫细胞化学(ICC) (图 1) 和免疫组织化学(IHC / IHC-P) (图 2)中的微管网络。
针对α-微管蛋白,Synaptic Systems提供广泛种类的单克隆,多克隆和重组抗体。并且不区分不同微管蛋白的翻译后修饰。
使用我们的α-微管蛋白抗体,并选择结合其它抗体,将在多种实验中给你最大的自由。
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| 302 008 | α-Tubulin, rabbit, monoclonal, recombinant IgGrecombinant IgG | WB ICC IHC | 100 µg | US$420.00 | |
| 302 203 | α-Tubulin, rabbit, polyclonal, affinity purifiedaffinity | WB ICC IHC ELISA | 100 µg | US$380.00 | |
| 302 206 | α-Tubulin, chicken, polyclonal, affinity purifiedaffinity | WB ICC IHC IHC-P | 50 µg | US$385.00 | |
| 302 211 | α-Tubulin, mouse, monoclonal, recombinant IgGrecombinant IgG | WB IP ICC IHC IHC-P ExM ELISA | 100 µg | US$420.00 | |
| 302 211C3 | α-Tubulin, mouse, monoclonal, purified IgG IgG, Oyster 550 discontinued | ICC | 50 µg | ||
| 302 217 | α-Tubulin, rat, monoclonal, purified IgG IgG | WB ICC IHC | 100 µg | US$420.00 | |
| 302 308 | α-Tubulin, Guinea pig, monoclonal, recombinant IgGrecombinant IgG | WB ICC IHC IHC-P | 50 µg | US$420.00 | |
| 302 411 | α-Tubulin, mouse, monoclonal, purified IgM IgM K.O. | WB ICC IHC | 100 µg | US$420.00 | |
| 302 204 | α-Tubulin , Guinea pig, polyclonal, antiserumantiserum | WB ICC IHC | 100 µl | US$370.00 |
通常,微管的半衰期很短,仅为5-10分钟。然而,一些翻译后修饰微管蛋白变体的半衰期可以持续数小时。总之,翻译后修饰可调节微管蛋白的聚合和解聚,在细胞骨架的动态微调中扮演着重要的角色(Infante, 2000)。
去酪氨酸化 /酪氨酸化:α-微管蛋白的C端络氨酸是由基因编码的,但是可以被血管抑制蛋白翻译后移除。从而使新创C端的一个自由谷氨酸暴露(Aillaud et al., 2017)。这个微管蛋白变体经常被当作Glu-微管蛋白,可在例如神经元或肾小球中找到(图 3)。这种去酪氨酸化反应可被微管络氨酸连接酶(TTL)逆转。酪氨酸化的α-微管蛋白被称为Tyr-微管蛋白,对新生小鼠胚胎的存活至关重要(Erck et al. 2005)。
多谷氨酰化: 数个谷氨酸(通常为 4-6个)被添加在位于α和β微管蛋白c端的谷氨酸残基中(Edde et al., 1990; Wehland et al., 1992)。这种多谷氨酰化的微管蛋白发生在例如神经元细胞和有丝分裂纺锤体中。
Synaptic Systems 为Glu-微管蛋白,多谷氨酰化微管蛋白及Tyr-微管蛋白提供成熟优质的单克隆抗体。
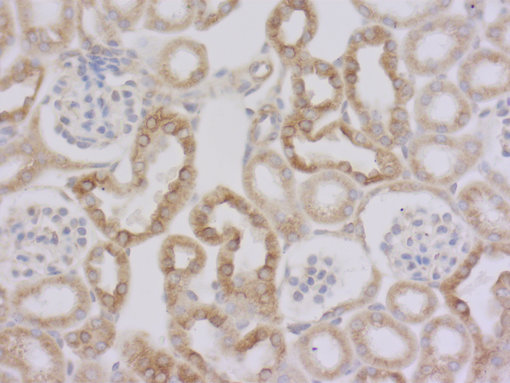
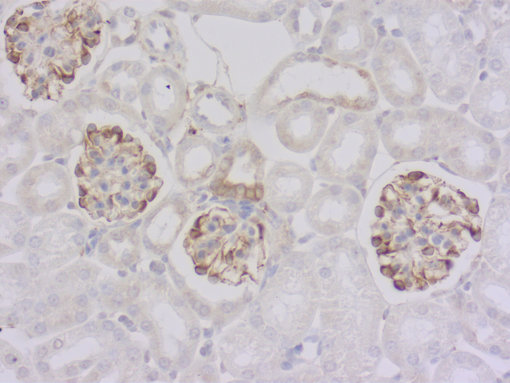
图 3a: 使用大鼠抗Tyr-α微管蛋白抗体对经PFA固定,石蜡包埋的小鼠肾切片进行间接免疫染色。(左, cat. no. 302 117, 稀释比例 1 : 1000; DAB)。
图 3b: 鼠抗 Glu-α微管蛋白抗体(cat. no. 302 011, 稀释比例 1 : 1000; DAB)。细胞核被苏木精染色可见为蓝色。
Delta2: α-微管蛋白最后两个残基的不可逆去除导致delta2或者D2-微管蛋白的形成。这种高度稳定的α -微管蛋白变体局限于神经元细胞,所以通常用来标记神经元细胞骨架的标记物 (Paturle-Lafanechère et al., 1994) (图 4)。
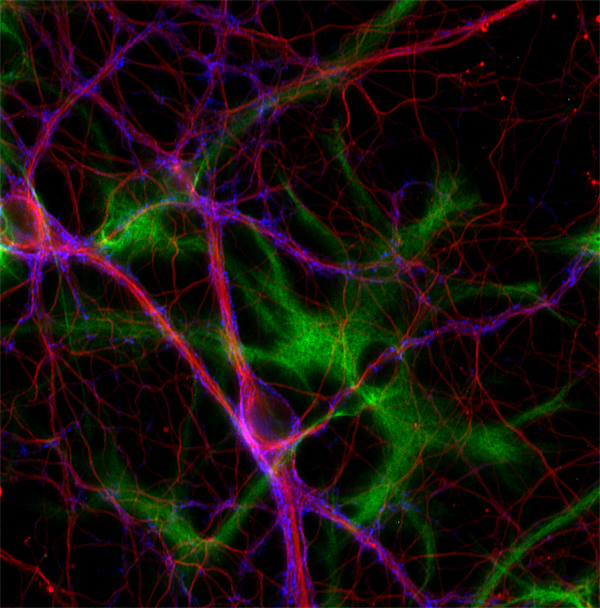
图4: 使用抗Δ2-微管蛋白 (cat. no. 302 213, 稀释比例 1 : 500; 红色)抗体,小鼠抗突触结合蛋白1 (cat. no. 105 011, 稀释比例 1 : 500; 蓝色) 和 豚鼠抗GFAP (cat. no. 173 004, 稀释比例 1 : 500; 绿色)抗体,对大鼠海马体神经元细胞和星形胶质细胞进行间接免疫染色。
β3-管蛋白
第三类β-管蛋白也被称为TuJ1,在中枢和外周神经系统(CNS和PNS)中含量丰富,在胎儿和出生后的发育过程中表达突出(Katsetos et al.,2003。据报道,它也在某些非神经元来源的肿瘤中表达(Dráberovà et al.,1998)(图5。
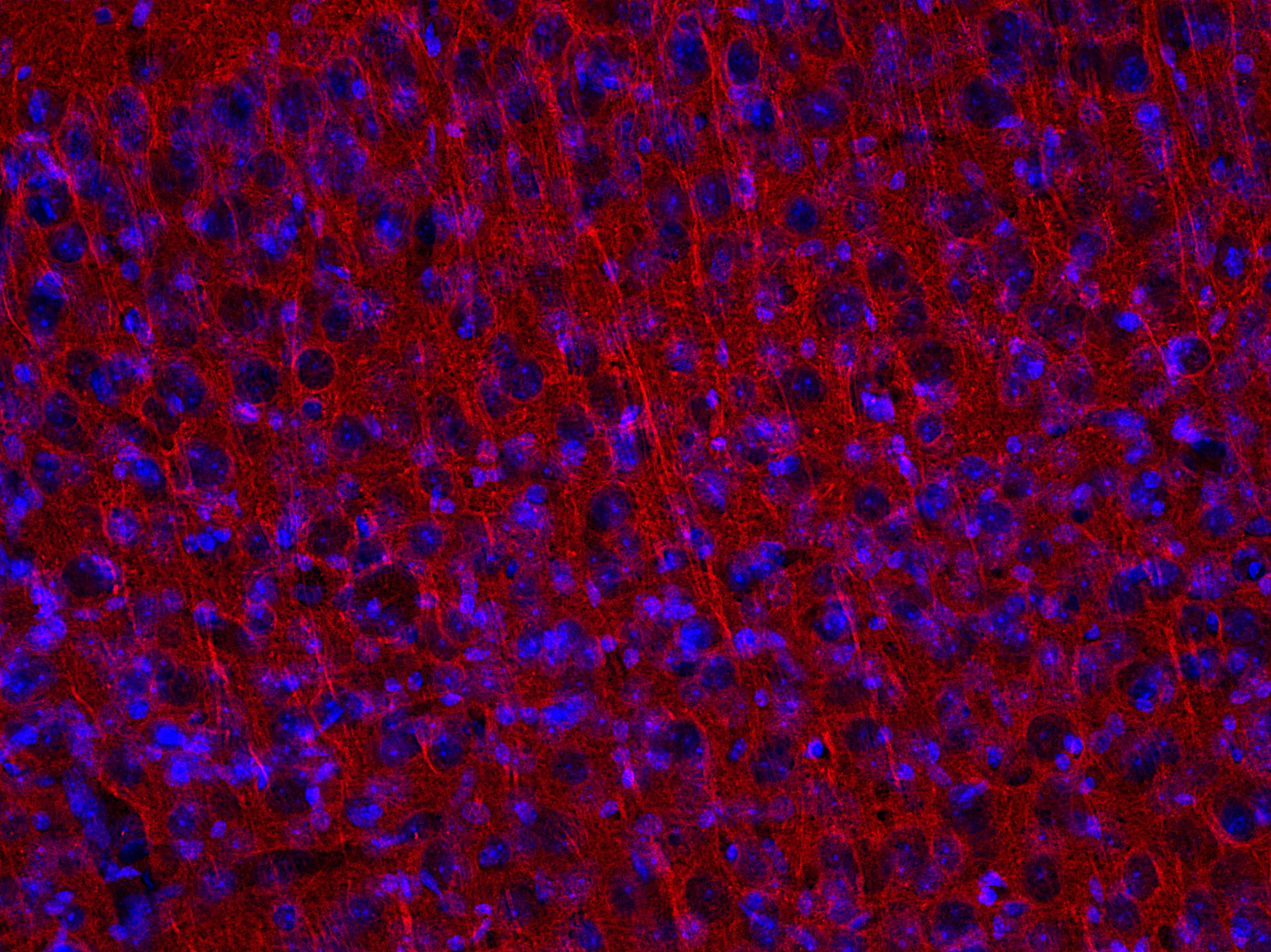
图 5: 使用小鼠抗β3-微管蛋白 (cat. no. 302 311, 稀释比例 1 : 5000, 红色)抗体,对经PFA固定的小鼠大脑皮层切片进行间接免疫染色。细胞核由DAPI染色可见为蓝色
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| 302 302 | β3-Tubulin, rabbit, polyclonal, antiserumantiserum | WB IP ICC IHC IHC-P | 200 µl | US$365.00 | |
| 302 304 | β3-Tubulin, Guinea pig, polyclonal, antiserumantiserum | WB IP ICC IHC IHC-P | 100 µl | US$375.00 | |
| 302 306 | β3-Tubulin, chicken, polyclonal, IgY fractionIgY fraction | WB ICC IHC IHC-P | 200 µl | US$365.00 | |
| 302 311 | β3-Tubulin, mouse, monoclonal, purified IgG IgG | WB IP ICC IHC IHC-P | 100 µg | US$420.00 | |
| 302 213 | Δ2-tubulin, rabbit, polyclonal, affinity purifiedaffinity | WB ICC IHC IHC-P | 50 µg | US$380.00 | |
| 302 011 | Glu-Tubulin, mouse, monoclonal, purified IgG IgG | WB ICC IHC IHC-P | 100 µg | US$420.00 | |
| 302 117 | Tyr-α-tubulin, rat, monoclonal, purified IgG IgG K.O. | WB IP ICC IHC IHC-P | 100 µg | US$420.00 |
Aillaud et al., 2017: Vasohibins/SVBP are tubulin carboxypeptidases (TCPs) that regulate neuron differentiation. PMID: 29146868
Dráberovà et al., 1998: Expression of class III beta-tubulin in normal and neoplastic human tissues. PMID: 9541471
Edde et al., 1990: Posttranslational glutamylation of alpha-tubulin. PMID: 1967194
Erck et al., 2005: A vital role of tubulin-tyrosine-ligase for neuronal organization. PMID: 15899979
Fletcher and Mullins, 2010: Cell mechanics and the cytoskeleton. PMID: 20110992
Goodson and Jonasson, 2018: Microtubules and Microtubule-Associated Proteins. PMID: 29858272
Infante, 2000: Detyrosinated (Glu) microtubules are stabilized by an ATP-sensitive plus-end cap. PMID: 11058078
Katsetos et al. 2003: Class III beta-tubulin in human development and cancer. PMID: 12740870
Paturle-Lafanechère et al., 1994: Accumulation of delta 2-tubulin, a major tubulin variant that cannot be tyrosinated, in neuronal tissues and in stable microtubule assemblies. PMID: 7962195
Wehland et al., 1992: Class II tubulin, the major brain β tubulin isotype is polyglutamylated on glutamic acid residue. PMID: 1379548
Certificates
ISO 9001 2015 Quality Management System and Green Lab Platinum certification level for sustaining laboratory processes.


Newsletter
Sign up for our newsletter and get the latest updates and news.