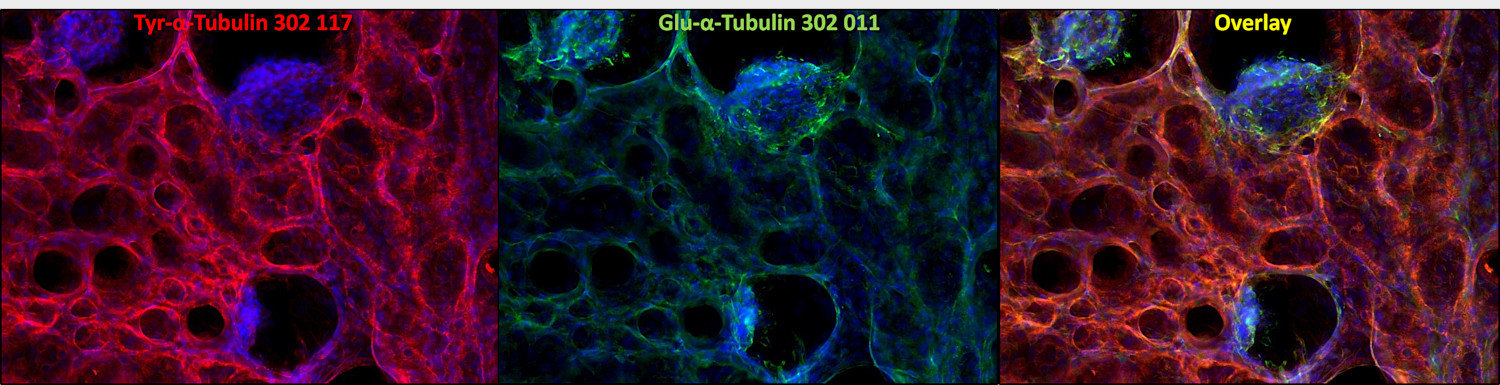
Cat. No.: 302 011
Amount: 100 µg
Price:
$420.00
|
|

|
|
| Cat. No. 302 011 |
100 µg purified IgG, lyophilized. Albumin and azide were added for stabilization. For reconstitution add 100 µl H2O to get a 1mg/ml solution in PBS. Then aliquot and store at -20°C to -80°C until use. Antibodies should be stored at +4°C when still lyophilized. Do not freeze! |
| Applications | |
| Clone | 1D5 |
| Subtype | IgG1 (κ light chain) |
| Immunogen | Synthetic peptide corresponding to residues near the c-terminus of human Glu-α-tubulin. (UniProt Id: Q71U36) |
| Epitop |
AA 448 to 450 from human Glu-α-tubulin (UniProt Id: Q71U36) |
| Reactivity |
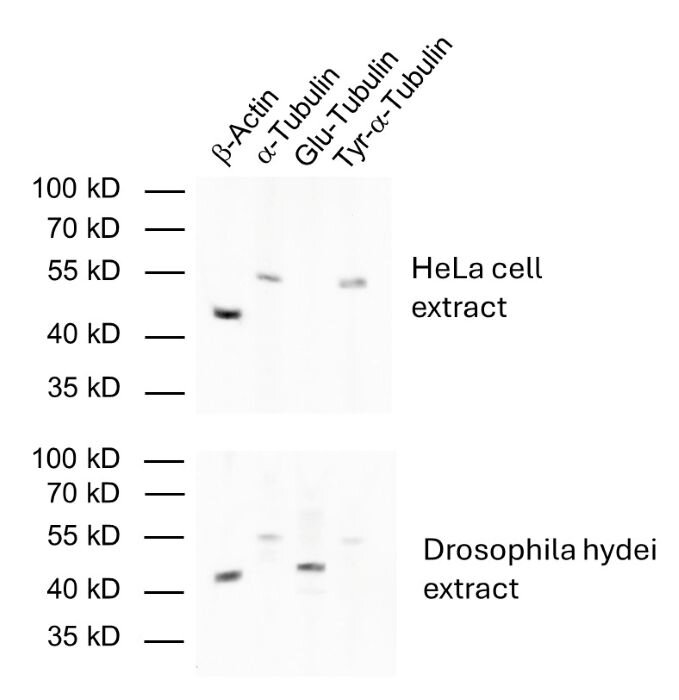
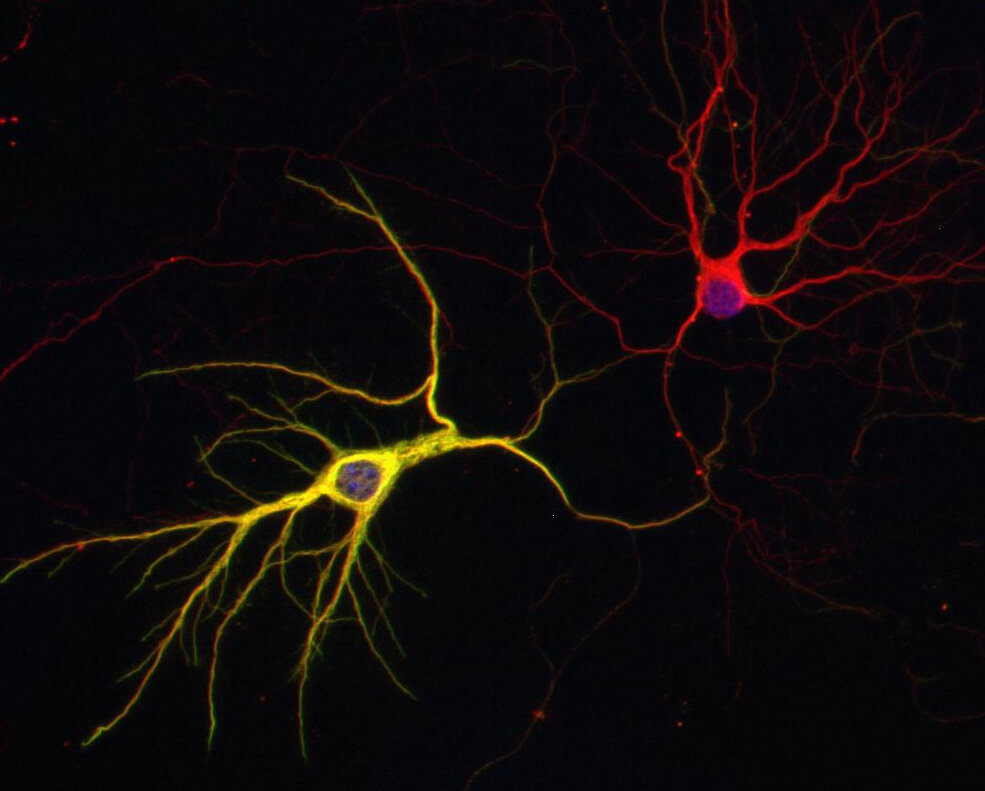
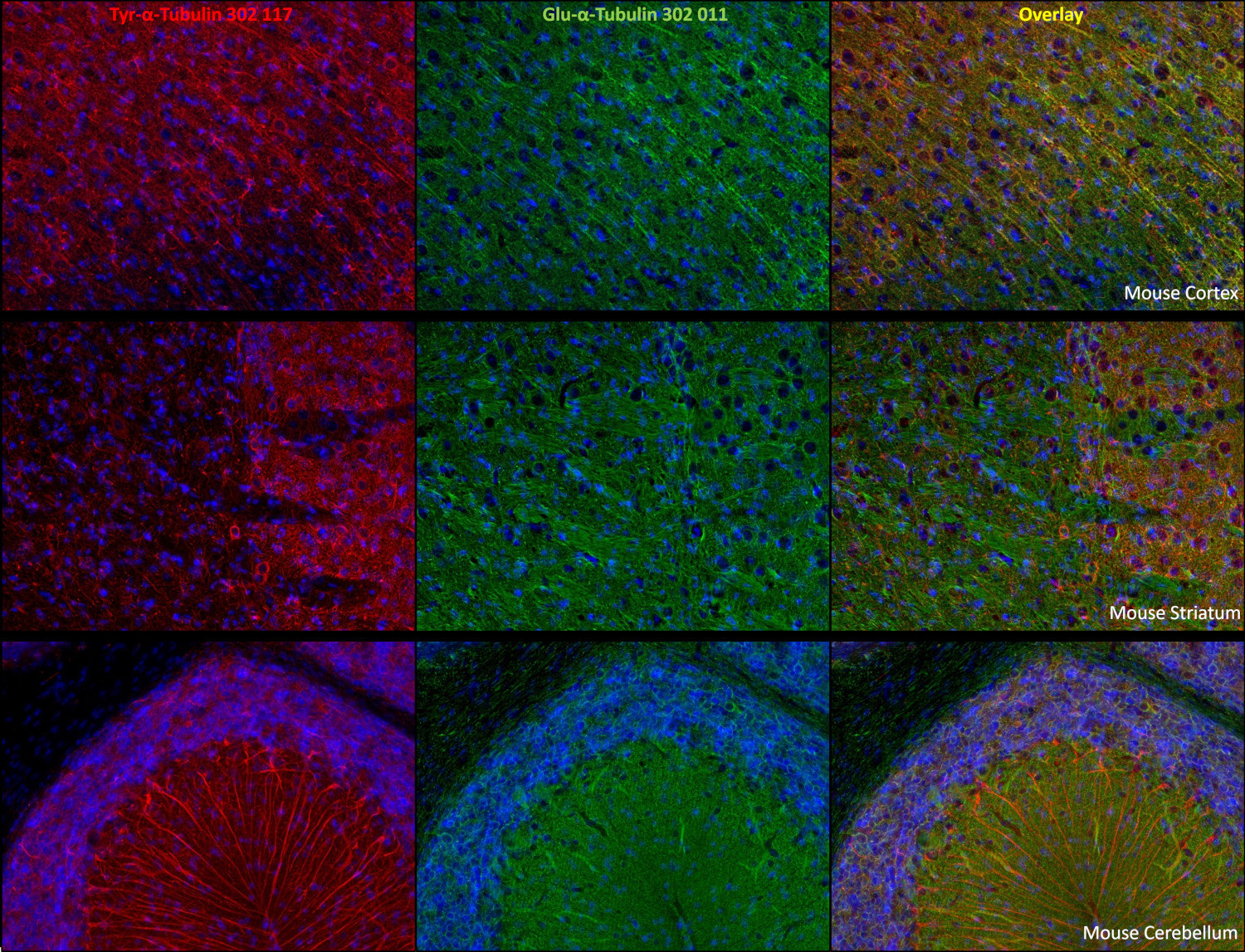
Reacts with: human (Q71U36), rat (P68370), mouse (P68369), zebrafish, eukaryotes, other vertebrates, Drosophila melanogaster. Other species not tested yet. Detects also cilia of Paramecium. |
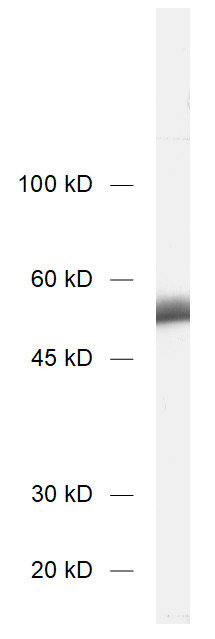
| Specificity | Specific for detyrosinated α-tubulin (glu-tubulin) and polyglutamylated tubulin (also β-tubulin). No cross reaction to tyrosinated tubulin. |
| Data sheet | Datasheet 302_011 |
|
|
Microtubules are involved in a wide variety of intracellular events including cell division, intracellular transport and secretion, axonal transport, and maintenance of cell morphology. They are composed of tubulin, a heterodimeric protein, consisting of two polypeptides, α-tubulin and β-tubulin (1).
α Tubulin undergoes numerous post-translational modifications that include tyrosination-detyrosination and deglutamylation, phosphorylation, acetylation, polyglutamylation, and polyglycylation. In one of the major posttranslational modifications, the C-terminal tyrosine residue in α-tubulin is added or removed reversibly, producing Glu-tubulin (after detyrosination) and Tyr-tubulin (with re-added tyrosine). Early stages of cell development are often enriched in Tyr tubulin, whereas mature cells show increased Glu tubulin in stable structures. Some microtubule associated proteins (MAPs), motor proteins like kinesins, or stabilizing factors have different affinities for Glu- or Tyr-tubulin (2,3,4).
A third variant of detyrosinated α-tubulin is Δ2-tubulin which lacks the C-terminal glutamic acid. It cannot be tyrosinated by tyrosine ligase and is one of the dominant α-tubulin isoforms in neurons (5).
Certificates
ISO 9001 2015 Quality Management System and Green Lab Platinum certification level for sustaining laboratory processes.


Newsletter
Sign up for our newsletter and get the latest updates and news.