-
|
-
• 简介
• 谷氨酸和阴离子的转运
• EAATs的分布
• 产品
• 参考文献
谷氨酸是大脑中最重要的兴奋型神经递质。许多不同的离子型和代谢型受体可介导其刺激信号到神经元。但是,在开关中,“关”信号和“开”信号一样重要。兴奋型氨基酸转运蛋白家族(EAATs)可通过将谷氨酸再循环回神经元和星形胶质细胞,迅速将谷氨酸从突触间隙和周围的细胞外空间中移除,从而协助终止谷氨酸的兴奋型信号。通过这种方式,EAATs可做到:
• 维持谷氨酸能信号的时间保真度。通过移除突触间隙中的谷氨酸,可以防止初始信号发出后突触后受体受到过度刺激,从而影响对下一个信号到达的检测。
• 维持信号的空间保真度,防止谷氨酸从已释放到的突触中“逃逸”,刺激非预期的突触外或突触谷氨酸受体
• 保护神经元免受细胞外空间过量谷氨酸的毒性作用。谷氨酸受体的激活加剧或延长会引发一系列神经毒性反应,最终导致神经元功能丧失和细胞死亡,这被称为兴奋性毒性
EAATs作为同质转运体,可将一个K+离子转运出细胞,同时将一个谷氨酸神经递质/分子(天冬氨酸也可以运输)和三个Na+离子以及一个H+离子交换进细胞中 (Alleva et al., 2022),见图1(改编自Freidman et al., 2020)。这种转运依赖于钠离子的电化学梯度,并由细胞膜上EAATs的同三聚体或异三聚体来促进(Kovermann et al., 2022)。
EAATs不仅仅是谷氨酸转运体,也是阴离子通道(Cl+),并在响应谷氨酸转运周期的转变中开放(Otis & Jahr, 1998)。
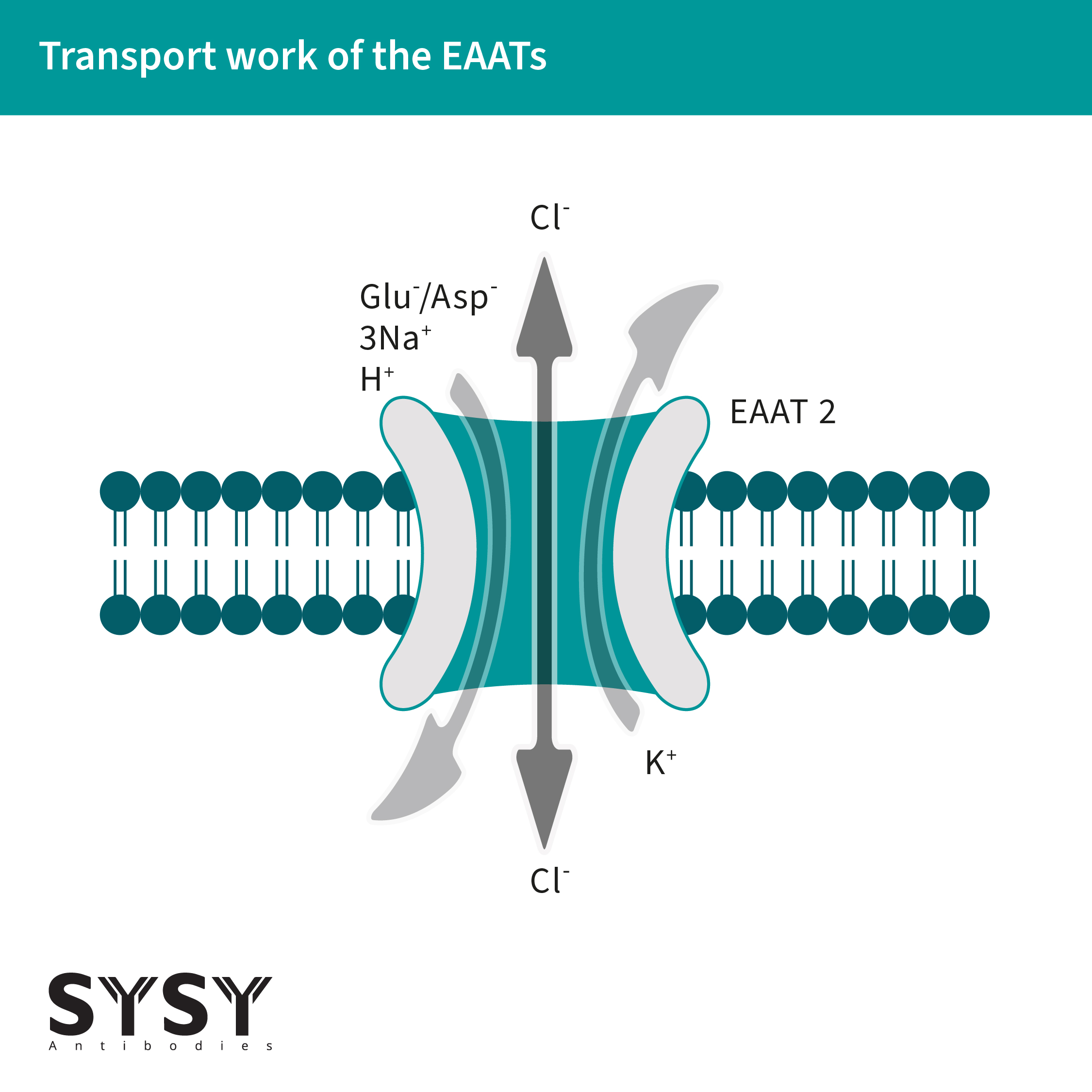
图 1: 谷氨酸和阴离子可通过 EAATs进行转运。
最近发表的数据表明,EAAT5,EAAT家族的一个视网膜特异性成员,不仅可作为谷氨酸进口商,也可以作为谷氨酸门控的Cl-通道,尤其在视锥光感受器中(Lukasiewcz et al., 2021)。
当谷氨酸被EAATs吸收到胶质细胞中时,会被转化为谷氨酰胺,随后被运输回突触前神经元,再转化为谷氨酸,并在VGLUTs的作用下被吸收到突触囊泡中。这一过程被命名为谷氨酸-谷氨酰胺循环(Andersen & Schousboe, 2023),见图2。
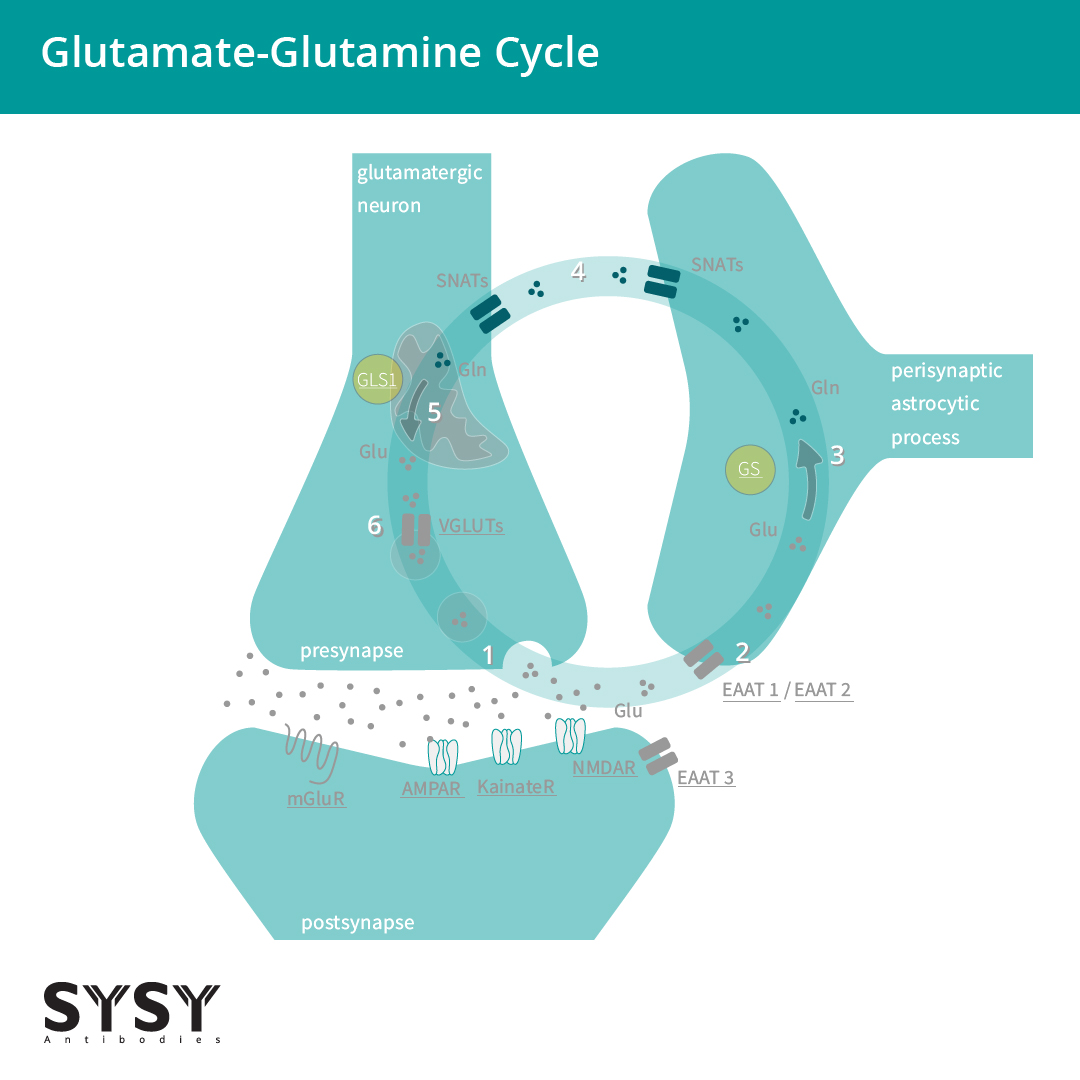
图 2: 谷氨酸能神经元和星形胶质细胞之间的谷氨酸-谷氨酰胺循环。
(1)谷氨酸(Glu)被释放并与亲离子受体和代谢型受体结合(AMPAR/GluA,KainateR/GluK,NMDAR/GluN,mGluRs)。(2)谷氨酸主要被星形胶质细胞通过兴奋型氨基酸转运体 EAAT 1/2吸收,部分可被神经元通过 EAAT3吸收。(3)谷氨酸(Glu)由星形细胞谷氨酰胺合成酶转化为谷氨酰胺(Gln)。(4)突触惰性谷氨酰胺从星形胶质细胞转移到神经元。(5)谷氨酰胺(Gln)由线粒体glutaminase 谷氨酰胺酶1 (GLS1)转化回谷氨酸(Glu)。(6)谷氨酸通过囊泡谷氨酸转运蛋白(VGLUTs)转运到囊泡中,并为下一轮转运做准备。
EAAT家族目前由EAAT1至EAAT5五个成员组成,它们具有各自不同的谷氨酸摄取动力学和不同程度的氯离子渗透和分布(Todd & Harding, 2020),见表1。
表1:
|
EAAT1:
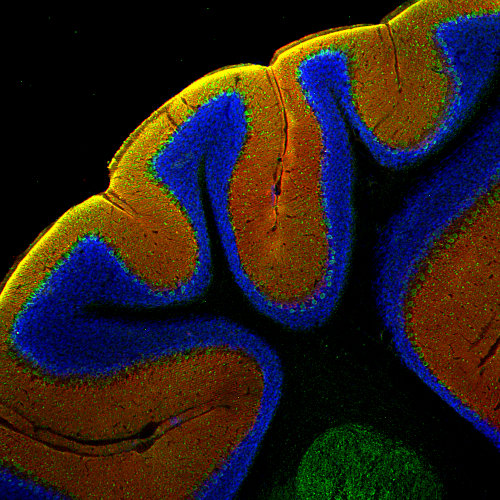
EAAT1也被称为GLAST-1,可在整个中枢神经系统中表达,并在小脑中星形胶质细胞和伯格曼胶质细胞中高度表达。在视网膜中,EAAT1在米勒细胞中表达。
强大的EAAT表达,特别是EAAT1,是一个广泛使用的成体神经干细胞(NSC)表型标记(Rieskamp et al., 2023)。
EAAT1,我们提供几种经敲除验证的抗体用于检测EAAT1。
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| 250 103 | EAAT1, rabbit, polyclonal, affinity purifiedaffinity K.O. extracellular | WB | 50 µg | US$380.00 | |
| 250 113 | EAAT1, rabbit, polyclonal, affinity purifiedaffinity K.O. cytoplasmic domain | WB IP ICC IHC IHC-P (FFPE) | 50 µg | US$380.00 | |
| 250 114 | EAAT1, Guinea pig, polyclonal, antiserumantiserum cytoplasmic domain | WB ICC IHC IHC-P (FFPE) | 100 µl | US$370.00 | |
| 250 116 | EAAT1, chicken, polyclonal, IgY fractionIgY fraction cytoplasmic domain | WB ICC IHC | 200 µl | US$365.00 | |
| 250-11P | EAAT1, control peptidecontrol peptide cytoplasmic domain | 100 µg | US$110.00 | ||
| 250-1P | EAAT1, control peptidecontrol peptide extracellular | 100 µg | US$110.00 |
EAAT2:
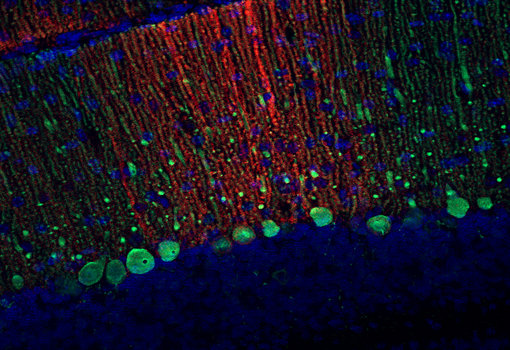
EAAT2,也被称为GLT-1,是大脑中最丰富的EAATs。它主要位于星形细胞分支,可分别在小脑和海马中高度表达(Yeung et al., 2021)。最近,在几种神经退行性疾病,如阿尔茨海默病、多发性硬化症和肌萎缩侧索硬化症(ALS)中,Dahlmanns等人综述发现了EAAT2的减少。在这种情况下,值得注意的是,早老素1 (PS1), γ分泌酶的活性亚基,可直接与EAAT2相互作用并影响转运体的细胞表面定位(Perrin et al., 2024)。
对于检测EAAT2,我们可提供优良的、经敲除验证的兔和豚鼠多克隆抗体。此外,一种小鼠单克隆抗体在WB、ICC、IHC和FFPE (IHC- p)应用中显示出了优异的结果。
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| 250 203 | EAAT2, rabbit, polyclonal, affinity purifiedaffinity K.O. extracellular | WB ICC IHC IHC-P (FFPE) | 50 µg | US$385.00 | |
| 250 204 | EAAT2, Guinea pig, polyclonal, antiserumantiserum K.O. extracellular | WB ICC IHC IHC-P (FFPE) | 100 µl | US$370.00 | |
| 250 208 | EAAT2, rabbit, monoclonal, recombinant IgGrecombinant IgG extracellular | WB ICC IHC IHC-P (FFPE) | 50 µg | US$420.00 | |
| 250 211 | EAAT2, mouse, monoclonal, purified IgG IgG extracellular | WB ICC IHC IHC-P (FFPE) | 100 µg | US$420.00 | |
| 250-2P | EAAT2, control peptidecontrol peptide extracellular | 100 µg | US$110.00 |
EAAT3:
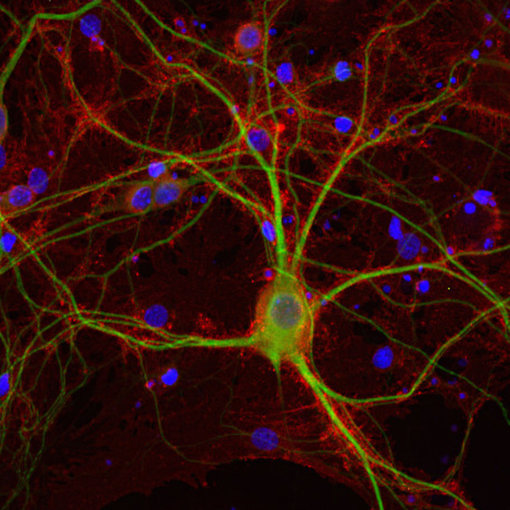
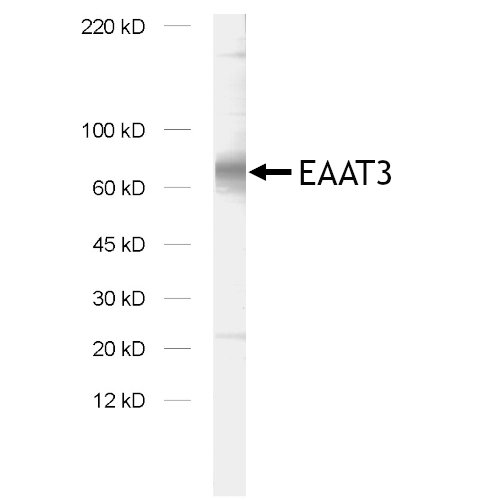
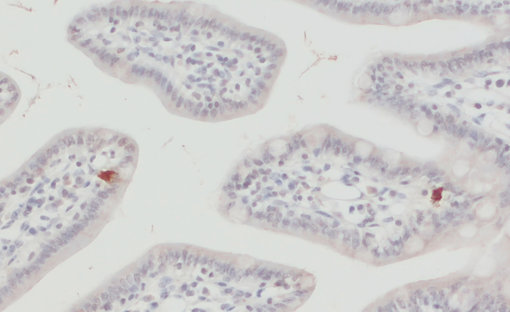
EAAT3,也被称为EAAC1或SLC1A1,是大脑中的“神经元”谷氨酸转运蛋白,主要位于大脑皮层、海马体、纹状体和基底神经节的轴突末梢和树突(Escobar et al., 2019)。在Escobar等人综述中,最新的研究结果假设EAAT3表达与强迫症(OCD)之间存在联系。在中枢神经系统外,EAAT3也可在肺、小肠、骨骼肌、肾外髓、髓线和皮质中发现(Todd & Hardingham, 2020)。
在我们的系列产品中,我们提供2种优越的兔多克隆抗体。
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| 250 303 | EAAT3, rabbit, polyclonal, affinity purifiedaffinity currently not available cytoplasmic domain | ICC IHC | 50 µg | US$380.00 | |
| 250 313 | EAAT3, rabbit, polyclonal, affinity purifiedaffinity cytoplasmic domain | WB | 50 µg | US$380.00 | |
| 250-31P | EAAT3, control peptidecontrol peptide cytoplasmic domain | 100 µg | US$110.00 |
EAAT4:
EAAT4,也被称为SLC1A6,是一个主要的神经元谷氨酸进口商。它主要位于小脑的浦肯野细胞,在前脑和中脑的某些亚区有少量表达(Massie et al., 2008)。最近的数据显示,EAAT4的表达遵循与醛缩酶C(zebrin)表达相似的旁矢状带模式,形成具有高水平和低水平EAAT4的分子多样性浦肯野细胞微区(Malhotra et al., 2021)。与EAAT1和EAAT2相比,EAAT4的谷氨酸转运能力较低,因此有研究者假设EAAT4在生理上主要作为阴离子通道(Suslova et al., 2023)。
在我们的新系列产品中,现在有2种优秀的兔抗EAAT4多克隆抗体。
EAAT5:
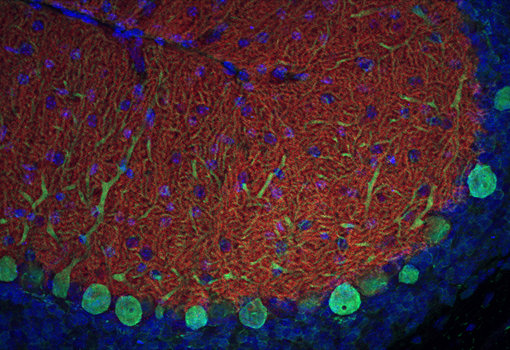
EAAT5蛋白,有时也被称为AAAT,主要存在于脊椎动物视网膜中,在那里它(与EAAT2一起)将谷氨酸转运到杆状细胞、锥体细胞和杆状双极细胞中。EAAT5转运蛋白聚集在突触带下方,位于突触前杆状细胞和锥体细胞的活跃区,完美的定位使得它可以捕捉到最近释放的谷氨酸。EAAT5的抑制表明该蛋白对视网膜中甘氨酸能无长突细胞(所有无长突细胞)的时间信号分辨率很重要(Tang et al., 2022)。在中枢神经系统外,EAAT5也在肝、肾、肠、心、肺和肌肉中表达。我们提供针对EAAT5的多克隆豚鼠经敲除验证的抗体。
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| 250 504 | EAAT5, Guinea pig, polyclonal, antiserumantiserum | IHC IHC-P (FFPE) | 100 µl | US$370.00 |
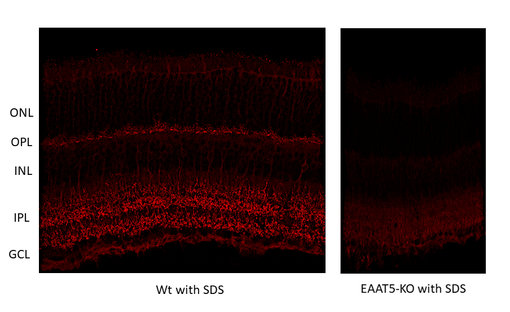
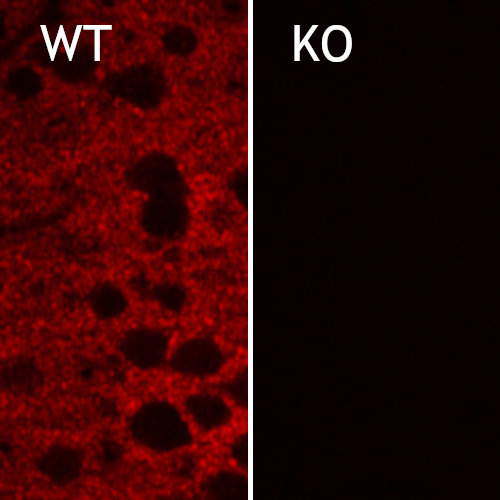
图10:对野生型(WT)和敲除型(KO)小鼠(cat. no. 250 504,稀释比例1:2000; red)视网膜中EAAT5进行间接免疫染色。根据(Gehlen et al. 2021),用4%甲醛浸泡固定组织,用1% SDS提取抗原。在此感谢来自德国于利希研究中心,分子和细胞生理学研究所,生物信息处理机构的Frank Müller和Christoph Aretzweiler-von Schwartzenberg友情提供图片。
图10:对野生型(WT)和敲除型(KO)小鼠(cat. no. 250 504,稀释比例1:2000; red)视网膜中EAAT5进行间接免疫染色。根据(Gehlen et al. 2021),用4%甲醛浸泡固定组织,用1% SDS提取抗原。在此感谢来自德国于利希研究中心,分子和细胞生理学研究所,生物信息处理机构的Frank Müller和Christoph Aretzweiler-von Schwartzenberg友情提供图片。
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| 250 103 | EAAT1, rabbit, polyclonal, affinity purifiedaffinity K.O. extracellular | WB | 50 µg | US$380.00 | |
| 250 113 | EAAT1, rabbit, polyclonal, affinity purifiedaffinity K.O. cytoplasmic domain | WB IP ICC IHC IHC-P (FFPE) | 50 µg | US$380.00 | |
| 250 114 | EAAT1, Guinea pig, polyclonal, antiserumantiserum cytoplasmic domain | WB ICC IHC IHC-P (FFPE) | 100 µl | US$370.00 | |
| 250 116 | EAAT1, chicken, polyclonal, IgY fractionIgY fraction cytoplasmic domain | WB ICC IHC | 200 µl | US$365.00 | |
| 250-11P | EAAT1, control peptidecontrol peptide cytoplasmic domain | 100 µg | US$110.00 | ||
| 250-1P | EAAT1, control peptidecontrol peptide extracellular | 100 µg | US$110.00 | ||
| 250 203 | EAAT2, rabbit, polyclonal, affinity purifiedaffinity K.O. extracellular | WB ICC IHC IHC-P (FFPE) | 50 µg | US$385.00 | |
| 250 204 | EAAT2, Guinea pig, polyclonal, antiserumantiserum K.O. extracellular | WB ICC IHC IHC-P (FFPE) | 100 µl | US$370.00 | |
| 250 208 | EAAT2, rabbit, monoclonal, recombinant IgGrecombinant IgG extracellular | WB ICC IHC IHC-P (FFPE) | 50 µg | US$420.00 | |
| 250 211 | EAAT2, mouse, monoclonal, purified IgG IgG extracellular | WB ICC IHC IHC-P (FFPE) | 100 µg | US$420.00 | |
| 250-2P | EAAT2, control peptidecontrol peptide extracellular | 100 µg | US$110.00 | ||
| 250 303 | EAAT3, rabbit, polyclonal, affinity purifiedaffinity currently not available cytoplasmic domain | ICC IHC | 50 µg | US$380.00 | |
| 250 313 | EAAT3, rabbit, polyclonal, affinity purifiedaffinity cytoplasmic domain | WB | 50 µg | US$380.00 | |
| 250-31P | EAAT3, control peptidecontrol peptide cytoplasmic domain | 100 µg | US$110.00 | ||
| 250 403 | EAAT4, rabbit, polyclonal, affinity purifiedaffinity | WB | 50 µg | US$385.00 | |
| 250 413 | EAAT4, rabbit, polyclonal, affinity purifiedaffinity | WB ICC IHC IHC-P (FFPE) | 50 µg | US$385.00 | |
| 250 504 | EAAT5, Guinea pig, polyclonal, antiserumantiserum | IHC IHC-P (FFPE) | 100 µl | US$370.00 | |
| End of List |
作者:Dr. Carsten Schmidt
Carsten 具有深厚的神经科学背景,多年来一直从事阿尔茨海默病的研究工作。

Alleva et al., 2022: Molecular Basis of Coupled Transport and Anion Conduction in Excitatory Amino Acid Transporters. PMID: 33587237
Andersen & Schousboe, 2023: Glial Glutamine Homeostasis in Health and Disease. PMID: 36322369
Dahlmanns et al., 2023: Glial Glutamate Transporter-Mediated Plasticity: System xc-/xCT/SLC7A11 and EAAT1/2 in Brain Diseases. PMID: 37005761
Escobar et al., 2019: The Neuronal Glutamate Transporter EAAT3 in Obsessive-Compulsive Disorder. PMID: 31803055
Freidman et al., 2020: Amino Acid Transporters and Exchangers from the SLC1A Family: Structure, Mechanism and Roles in Physiology and Cancer. PMID: 31981058
Kovermann et al., 2022: Cellular Physiology and Pathophysiology of EAAT Anion Channels. PMID: 35087380
Lukasiewcz et al., 2021: EAAT5 Glutamate Transporter-Mediated Inhibition in the Vertebrate Retina. PMID: 34025361
Malhotra et al., 2021: Climbing Fiber-Mediated Spillover Transmission to Interneurons Is Regulated by EAAT4. PMID: 34400517
Massie et al., 2008: High-affinity Na+/K+-dependent glutamate transporter EAAT4 is expressed throughout the rat fore- and midbrain. PMID: 18770868
Otis & Jahr, 1998: Anion currents and predicted glutamate flux through a neuronal glutamate transporter. PMID: 9736633
Perrin et al., 2024: Identification of PS1/gamma-secretase and glutamate transporter GLT-1 interaction site. PMID: 38499151
Rieskamp et al., 2023: Excitatory amino acid transporter 1 supports adult hippocampal neural stem cell self-renewal. PMID: 37534178
Suslova et al., 2023: Apo state pore opening as functional basis of increased EAAT anion channel activity in episodic ataxia 6. PMID: 37538371
Tang et al., 2022: Glutamate Transporters EAAT2 and EAAT5 Differentially Shape Synaptic Transmission from Rod Bipolar Cell Terminals. PMID: 35523583
Todd & Hardingham, 2020: The Regulation of Astrocytic Glutamate Transporters in Health and Neurodegenerative Diseases. PMID: 33348528
Yeung et al., 2021: EAAT2 Expression in the Hippocampus, Subiculum, Entorhinal Cortex and Superior Temporal Gyrus in Alzheimer’s Disease. PMID: 34588956
Certificates
ISO 9001 2015 Quality Management System and Green Lab Platinum certification level for sustaining laboratory processes.


Newsletter
Sign up for our newsletter and get the latest updates and news.
 and knockout (KO) animals with rabbit polyclonal anti-EAAT1")